得獎人:
Julius Axelrod 朱裏亞斯.阿庫賽爾洛德
Ulf Svante von Euler烏魯夫.S.旺.歐易勒
Bernhard Katz巴納德.卡茲
得獎內容:
發現了神經末梢中的體液性傳導物質及其貯存、釋放和失活的機制,
神經性衝動是電位的變化,它沿著神經快速的傳到終端,並使別的神經產生新的衝動,或引起肌肉、分泌線或生體其他器官動作器的活動。神經衝動的傳達,從以前就順理成章的被認為是由物理的手段發生,也就像電流經過電線一樣。但是,在邁入20世紀,亨利.德爾與奧特.列威指出,衝動傳達可由化學方法發生。在神經末端,衝動使化學物質釋出,這又使下面的神經或受神經支配的組織發生電氣活動。如此,使神經末端與受神經支配的組織間的機能間隙得以接通。而這種神經傳導的化學媒介物的發現,導入了革命性的思想。而在這個快速發展的領域之中,也產生許多心的疑問,如這強而有力的傳導物質如何形成、儲藏而釋放?傳導物質怎麼出來,發揮作用,又於一秒更短的時間內消失?此外,還有什麼物質與此過程有關?而這三位得獎人為此領域存在的問題,各自都有特別的貢獻。
Bernhard Katz巴納德.卡茲博士,運動神經的衝動在運動神經終盤作用引起肌肉活動而發生的電氣現象,特別感興趣。具有電容器性質的肌肉特殊構造因神經衝動而帶電,接著因其帶電引發肌肉活動。卡茲發現『微小終盤電位』之存在,也因而指出,在運動神經與肌肉終盤之間,有傳達物質乙脢之小塊由神經終端釋出。
Ulf Svante von Euler烏魯夫.S.旺.歐易勒博士則對交感神經系特別感興趣。在早期便確定正腎上腺素為腎腺素性神經傳導物質。後來,他與同事故尼爾斯.埃克.希拉布共同證明,副腎上腺素在神經中合成,並儲藏於約一萬分之一豪米的顆粒中,並對有關這些顆粒的性質研究,貢獻很大。
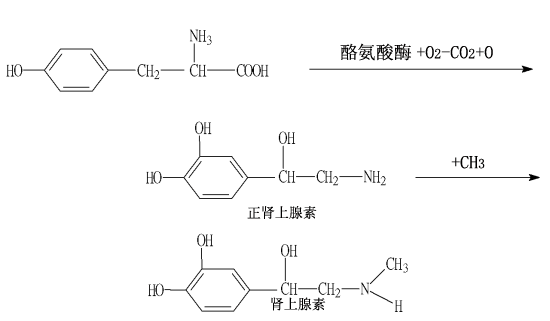
Julius Axelrod 朱裏亞斯.阿庫賽爾洛德博士,主要探討正腎上腺素由神經末端釋出後的路徑。由此他發現這傳導物質的甲基化引起的酵素不活性化,並對其性質加以研究。值得一提的是,他證明瞭由神經末梢再回收副腎上腺素的機構。神經傳導物質往往放出過多,立刻會大部分被回收到神經中端內的儲藏顆粒中。也就是說,這是限制神經衝動的作用時間,快速而有效的方法。
此三位博士所完成的,如此重要關於化學神經傳導過程之性質的基本研究,不但豐富了醫學理論領域的知識,對於神經末梢和中樞神經疾病的闡明和治療,也具有無法估計的重要性。
得獎者生平:

Ulf Svante von Euler烏魯夫.S.旺.歐易勒博士
Ulf.S.von.Euler 1905年2月7日生於斯德哥爾摩,他是Hans von Euler-Chelpin和Astrid
Cleve的次子。其父生於德國奧格斯堡,是Rigas von Euler-Chelpin將軍的獨子。Hans von Euler-Chelpin於1929年獲諾貝爾化學獎。歐易勒的外祖父Per Teodor Cleve是元素銩和鈥的發現者,烏普薩拉大學化學教授。歐易勒的母親Astrid Cleve曾獲得植物學的哲學博士學位,後來她大部分的科學活動都圍繞矽藻和地質學的研究,於1955年獲教授銜。
歐易勒先在斯德哥爾摩和卡爾斯塔德上學,1922年入卡羅琳學院學醫。他家庭中充滿學術氣氛,他又經常有機會與科學家接觸(1903年獲諾貝爾化學獎的Svante
Arrhenius是他的教父),這些無疑對他後來的熱衷於研究工作産生極大的影響。他的父母推動(但不強制)他進行研究工作。跟隨Robin
Fahraeus開始進行了一些自己的研究工作,並因對發熱時的血液具有血管收縮作用這一研究而獲獎,這更加激勵他從事研究工作。
1926年起,他在G.Liljestrand的藥理系當助手,1930年他在那裏寫成了學位論文,當年他被任命爲藥理學助理教授,在Liljestrand的不斷幫助下,他交上了好運,得到了洛克菲勒基金會的資助在國外從事研究工作(1930-1931),得以與H.H.Dale在倫敦合作,與I.de
Burgh Daly在伯明罕合作。這個時期,他在生理學和藥理學方面進行多種內容的研究,這爲他以後的研究打下了其意義無法估價的良好基礎。非常幸運,他在腸浸出物中發現了一種生物活性因數-P物質,後來又與Dale實驗室的J.H.Gaddum進一步對該物質進行研究。回國以後,他的興趣很快轉到了這個方面,他接著又陸續發現了前列腺素和精囊素(1935)、呱啶(1942)及去甲腎上腺素(1946)。
在倫敦的A.V.Hill實驗室中,奧伊勒對物理學的疑難問題和研究方法已有一些認識(1934)。1938年他在倫敦又從G.L.Brown那兒學到神經肌肉傳遞的研究方法。1946-1947年,他在布宜諾斯艾利斯B.A.Houssay實驗室與E.Brarn.Menéndez合作研究有關內分泌學和實驗性腎性高血壓的許多方面的問題。
他在1930-1939年擔任助理教授,後被任命爲卡羅琳學院生理學教授,這職位他一直擔任到1971年。從卡羅琳學院舊址遷到斯德哥爾摩郊外新址的現代化實驗室之後,工作條件得到了改善。醫學研究理事會於1950年後給了他們慷慨的支援,這大大促進了他們的研究工作;支援還來自私人基金會、制藥工業以及美國的研究基金會。
1946年,歐易勒鑒定出腎上腺素能神經遞質就是去甲腎上腺素,以後他的大部分研究工作就一直以此爲課題。他的實驗室研究了去甲腎上腺素在神經和器官中的分佈情況及其在各種生理和病理情況下的排泄情況,同時還對該種物質作了定量分析。他又同後來的同事N.Å.Hillarp一起發現了這種遞質存貯於亞細胞微粒之中,這給研究指出了新的方向。1958年起,除神經傳遞過程外,許多涉及遞質吸收、儲存及從神經顆粒釋出的問題一直是他們研究的主要課題。爲數衆多的大學生、助理研究員和副研究員都參加過這些研究。
歐易勒於1953-1960年爲諾貝爾生理學或醫學獎評定委員會委員,1960-1965年爲該委員會的秘書。1965年,奧伊勒教授被任命爲諾貝爾基金會的董事長。
歐易勒教授于1961年受蓋爾德納獎(加拿大),1965年受雅爾獎(挪威),1967年受斯托弗獎(美國),1953年受卡爾.路德維希獎章(德國),1969年受斯米德貝格獎章(德國),1970年受小聖母獎章(義大利)。
歐易勒教授是斯德哥爾摩皇家科學院院士、哥本哈根皇家科學院院士、海牙利奧波德科學院院士、巴賽隆納皇家醫學科學院院士及美國哲學學會會員,也是美國內科醫師學院、美國心臟協會臨床心臟學委員會、瑞典內科醫師學會、義大利藥理學會、瑞典內分泌學會和航空醫學學會的榮譽會員。
他接受過烏梅亞大學、裏約熱內盧大學、第戎大學、根特大學、蒂賓根大學、布宜諾斯艾利斯大學、愛丁堡大學、馬德裏大學、古斯塔夫斯.阿道弗斯學院的榮譽博士學位。
1930-1957年歐易勒與J.Sodenstierna結爲夫婦。他們有四個孩子:Hans
Leo,是美國貝塞斯達國立衛生研究院的科學管理人員;Johan Christopher,斯德哥爾摩塞拉菲默醫院的麻醉師;Ursula katarina,斯德哥爾摩大學藝術史系文學士助理;Marie
Jane,澳大利亞墨爾本的化學工程師。
1958年,歐易勒與Dagmar Cronstedt女伯爵結爲夫婦。

Julius Axelrod 朱裏亞斯.阿庫賽爾洛德博士
Julius Axelrod 1912年5月30日生於紐約市。1933年在紐約市學院獲理學士學位。1941年在紐約大學獲文學碩士學位。1955年在喬治華盛頓大學獲博士學位。
1933~1935年,他在紐約大學醫學院細菌系實驗室任助教,1935~1946年他在工業衛生實驗室人化學師,1946~1949年任Goldwater紀念醫院、紐約大學第三研究部的研究助理,1949~1950年爲NIH國立心臟研究所化學藥理學部的助理化學師,1950~1958年爲NIH國立心臟研究所的化學師,並于1953年成爲高級化學師。1955年他被任命爲健康、教育和福利部,健康服務心理衛生局,國立心理衛生研究所臨床科學實驗室藥理部主任。
阿庫賽爾洛德曾接受下列榮譽和獎勵:國家科學基金會遊學獎(1958);德國藥理學通訊會員(1959);國際生理學會遊學獎(1961);紐約大學醫學院Otto.loewi紀念講座(1964);神經精神病協作研究會卓越研究獎(1965);費城內分泌學會Karl
E.Paschkis紀念講座(1966);Gairdner基金會獎(1967);國立衛生研究所講座(1967);南加利福尼亞大學Nathanson紀念講座(1968);喬治華盛頓大學卓越成就獎(1968);DHEW高級貢獻獎(1968);蒙特利爾大學Claude
Bernard教授職位(1969);DHEW卓越貢獻獎(1970);現代醫學雜誌卓越貢獻獎(1970)。
阿庫賽爾洛德博士是Sigma Xi,國際腦研究組織、美國化學學會、美國藥理學和實驗治療學學會,美國生物化學家學會及美國科學促進會的成員。1961年他爲美國神經心理藥理學院的研究員(1966~1969爲理事會理事)。
阿庫賽爾洛德博士是許多雜誌的編委會成員,其中有藥理學和實驗治療學雜誌、醫學化學雜誌、生命科學、迴圈研究、神經生物學雜誌、藥理學研究通訊、神經化學雜誌和國際心理生物學雜誌。
1970年他成爲國立心理衛生研究所藥理研究部成員。
阿庫賽爾洛德 1938年與Sally Taub結婚,他們有兩個孩子。

Bernhard Katz巴納德.卡茲
Bernard Katz 1911年3月26日出生於德國來比錫,是俄羅斯猶太人後裔。卡茲是Max Katz和Eugenie
Rabinowitz的獨生兒子。1921~1929年他在來比錫艾伯特高級中學受教育,1929~1934年在來比錫大學學醫,1933年得到生理學研究的Siegfried
Garten獎金,並在1934年獲醫學博士學位。
他在1935年2月離開德國,有倫敦大學學院的A.V.Hill的實驗室工作到1939年,1939年他獲得哲學博士(倫敦大學)及Beit紀念研究員職務。1942年被授予科學博士學位(倫敦大學)。
1939年卡茲作爲一個卡內基研究成員參加到澳大利亞悉尼醫院的埃克爾斯的實驗室。他與埃克爾斯和S.W.Kuffler在神經肌肉研究中合作。
1941年,他從澳大利亞回到倫敦大學學院,又到A.V.Hill的研究單位,爲研究助理指導和Henry Head主任研究員(由皇家學會委任)。1950年他爲生理學講師,1955年起他在倫敦大學任生物物理教授。
1952年卡茲被選爲皇家學會會員,1968年爲皇家醫生學院的成員。1968年爲丹麥皇家科學和文學院的外國院士;1969年爲Lincei國家科學院的外國院士及美國藝術和科學院的院士,1961年爲倫敦大學學院的成員。1965年獲Feldberg基金獎,1967年獲皇家醫生學院Baly勳章和皇家學會Copley勳章。1969年他被封爲爵士。他自1967年起一直是農業研究理事會理事,自1968年起爲皇家學會生物學秘書。
卡茲教授研究的主要領域有:神經和肌肉研究,特別是神經肌肉傳遞的物理-化學機制。
1945年卡茲與新南威爾士的Cremorne的Marguerite Penly結婚。他們有兩個孩子:David(1947年生),在倫敦學習藥理學和醫學;Jonathan(1950年生),在牛津彭布羅克學院學習古典文學。
相關內容:
一、神經與骨骼肌的關係
人體各種形式的運動,主要是靠一些肌細胞的收縮活動來完成的。例如,軀體的各種運動和呼吸動作由骨骼肌的收縮來完成;心臟的射血活動由心肌的收縮來完成;一些中空器官如胃腸、膀胱、子宮、血管等器官的運動,則由平滑肌的收縮來完成。不同肌肉組織在功能和結構上各有特點,但從分子水平來看,各種收縮活動都與細胞內所含的收縮蛋白質,主要與肌凝蛋白和肌纖蛋白的相互作用有關;收縮和舒張過程的控制,也有某些相似之處。本節以研究最充分的骨骼肌爲重點,說明肌細胞的收縮機制。
骨骼肌是體內最多的組織,約占體重的40%。在骨和關節的配合下,通過骨骼肌的收縮和舒張,完成人和高等動物的各種軀體運動。骨骼肌由大量成束的肌纖維組成,每條肌纖維就是一個肌細胞。成人肌纖維呈細長圓柱形,直徑約60
μm,長可達數毫米乃至數十釐米。在大多數肌肉中,肌束和肌纖維都呈平行排列,它們兩端都和由結締組織構成的腱相融合,後者附著在骨上,通常四肢的骨骼肌在附著點之間至少要跨過一個關節,通過肌肉的收縮和舒張,就可能引起肢體的屈曲和伸直。我們的生産勞動、各種體力活動等,都是許多骨骼肌相互配合的活動的結果。每個骨骼肌纖維都是一個獨立的功能和結構單位,它們至少接受一個運動神經末梢的支配,並且在體骨骼肌纖維只有在支配它們的神經纖維有神經衝動傳來時,才能進行收縮。因此,人體所有的骨骼肌活動,是在中樞神經系統的控制下完成的。
一、神經-骨骼肌接頭處的興奮傳遞
運動神經纖維在到達神經末梢處時先失去髓鞘,以裸露的軸突末梢嵌入到肌細胞膜上稱作終板的膜凹陷中,但軸突末梢的膜和終板膜並不直接接觸,而是被充滿了細胞外液的接頭間隙隔開,其中尚含有成分不明的基質;有時神經末梢下方的終板膜還有規則地再向細胞內凹入,形成許多皺褶,其意義可能在於增加接頭後膜的面積,使它可以容納較多數目的蛋白質分子,它們最初被稱爲N-型乙酰膽鹼受體,現已證明它們是一些化學門控通道,具有能與ACh特異性結合的亞單位。在軸突末梢的軸漿中,除了有許多線粒體外還含有大量直徑約50nm的無特殊構造的囊泡(圖2-19)。用組織化學的方法可以證明,囊泡內含有ACh;此ACh首先在軸漿中合成,然後貯存在囊泡內。據測定,每個囊泡中貯存的ACh量通常是相當恒定的,且當它們被釋放時,也是通過出胞作用,以囊泡爲單位“傾囊”釋放,被稱爲量子式釋放。在神經末梢處於安靜狀態時,一般只有少數囊泡隨機地進行釋放,不能對肌細胞産生顯著影響。但當神經末梢處有神經衝動傳來時,在動作電位造成的局部膜去極化的影響下,大量囊泡向軸突膜的內側面靠近,通過囊泡膜與軸突膜的融合,並在融合處出現裂口,使囊泡中的ACh全部進入接頭間隙。據推算,一次動作電位的到達,能使大約200~300個囊泡的內容排放,使近107個ACh分子被釋放。軸突末梢處的電位變化引起囊泡排放的過程十分複雜,但首先是軸突末梢膜的去極化,引起了該處特有的電壓門控式Ca2+通道開放,引起細胞間隙液中的Ca2+進入軸突末梢,觸發了囊泡移動以至排放的過程。Ca2+的進入量似乎決定著囊泡釋放的數目;細胞外液中低Ca2+或(和)高Mg2+,都可阻礙ACh的釋放而影響神經-肌接頭的正常功能。已故馮德培院士在30年代對神經-肌接頭的化學性質傳遞進行過重要的研究。
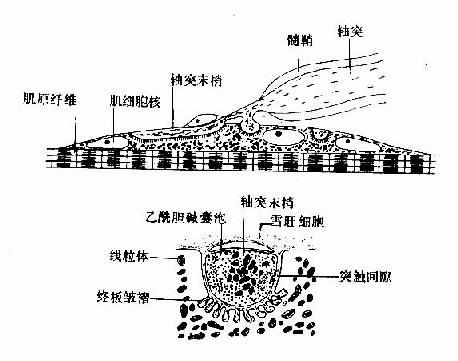
圖2-19 神經-肌接頭處的超微結構示意圖
當ACh分子通過接頭間隙到達終板膜表面時,立即同集中存在於該處的特殊通道蛋白質的兩個α-亞單位結合,每分子的通道將結合兩個分子的ACh,由此引起的蛋白質分子內部構象的變化會導致它的通道結構的開放。這種通道開放時,孔道的橫截面比前面提到的Na+通道的面積爲大,可允許Na+、K+甚至少量Ca2+同時通過;由於這幾種離子正常時在膜內處的分佈特點,實際出現的是Na+的內流和K+的外流,其總的結果是使終板膜處原有靜息電位減小,向零值靠近,亦即出現膜的去極化;這一電變化,稱爲終板電位,它的出現約較神經衝動到達接頭前膜處晚0.5~1.0ms。有人曾在運動神經無衝動到達末梢時,記錄到由於個別囊泡的自發釋放在終板膜上引起的微小的電變化,稱爲微終板電位。終板電位與前述的局部興奮電反應有類似的性質:不表現“全或無”特性,其大小與接頭前膜釋放的ACh的量成比例;無不應期,可表現總和現象等,如我們一般記錄到的終板電位就是多數微終板電位總和的結果。終板電位産生時,它將以電緊張性擴布的形式影響終板膜周圍一般肌細胞膜。一般的肌細胞膜與神經軸突的膜性質類似,其中主要含電壓門控式Na+通道和K+通道;因而當同終板膜鄰接的肌細胞膜的靜息電位由於終板電位的影響而去極化到該處膜的閾電位水平時,就會引發一次向整個肌細胞膜作“全或無”式傳導的動作電位,後者再通過所謂“興奮-收縮耦聯”,引起肌細胞出現一次機械收縮。
正常情況下,一次神經衝動所釋放的ACh以及它所引起的終板電位的大小,大約超過引起肌細胞膜動作電位所需閾值的3~4倍,因此神經肌接頭處的興奮傳遞通常是1對1的,亦即運動纖維每有一次神經衝動到達末梢,都能“可靠地”使肌細胞興奮一次,誘發一次收縮;這一點與將來要講的神經元之間的興奮傳遞有明顯不同(見第十章)。接頭傳遞能保持1對1的關係,還要靠每一次神經衝動所釋放的ACh能夠在它引起一次肌肉興奮後被迅速清除,否則它將持續作用於終板而使終板膜持續去極化,並影響下次到來的神經衝動的效應。已知,ACh的清除主要靠膽鹼酯酶的降解作用來完成,此酶主要分佈在接頭間隙中和接頭後膜上,它們大約可以在2.0ms的時間內將一次神經衝動所釋放的ACh清除掉。許多藥物可以作用於接頭傳遞過程中的不同階段,影響正常的接頭功能。例如,美洲箭毒和α-銀環蛇毒可以同ACh競爭終板膜的ACh受體亞單位,因而可以阻斷接頭傳遞而使肌肉失去收縮能力;有類似作用的藥物稱爲肌肉鬆弛劑;有機磷農藥和新斯的明對膽鹼酯酶有選擇性的抑制作用,可造成ACh在接頭和其他部位的大量積聚,引起種種中毒症狀。重症肌無力是由於體內骨骼肌終板處的ACh門控通道數量不足或功能障礙所引起。
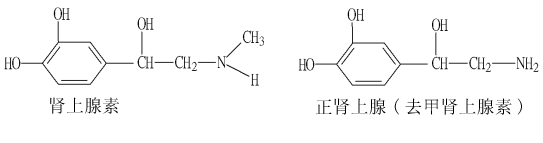
二、腎上腺素和正腎上腺素